Background: Stress cardiomyopathy (SCM) is a transient reversible left ventricular dysfunction that more often occurs in women. Symptoms of SCM patients are similar to those of acute coronary syndrome (ACS), but little is known about biomarkers. The goals of this study were to identify the potentially crucial genes and pathways associated with SCM.Methods: We analyzed microarray datasets GSE95368 derived from the Gene Expression Omnibus (GEO) database. Firstly, identify the differentially expressed genes (DEGs) between SCM patients in normal patients. Then, the DEGs were used for Gene Ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway enrichment analysis. Finally, the protein-protein interaction (PPI) network was constructed and Cytoscape was used to find the key genes.Results: In total, 25 DEGs were identified, including 10 upregulated genes and 15 downregulated genes. These DEGs were mainly enriched in ECM-receptor interaction, dilated cardiomyopathy (DCM), human papillomavirus infection, and focal adhesion, whereas in GO function classification, they were mainly enriched in the extracellular region, positive regulation of the multicellular organismal process, establishment of localization, and intracellular vesicle.Conclusion: Seven hub genes contained APOE, MFGE8, ALB, APOB, SAA1, A2M, and C3 identified as hub genes of SCM, which might be used as diagnostic biomarkers or molecular targets for the treatment of SCM.Pan XY, Zhang ZW. MFGE8, ALB, APOB, APOE, SAA1, A2M, and C3 as Novel Biomarkers for Stress Cardiomyopathy. Cardiovasc Ther. 2020;2020:1615826.
Background: A practical strategy to discover sepsis specific proteins may be to compare the plasma peptides and proteins from patients in the intensive care unit with and without sepsis. The aim was to discover proteins and/or peptides that show greater observation frequency and/or precursor intensity in sepsis. The endogenous tryptic peptides of ICU-Sepsis were compared to ICU Control, ovarian cancer, breast cancer, female normal, sepsis, heart attack, Alzheimer's and multiple sclerosis along with their institution-matched controls, female normals and normal samples collected directly onto ice.Methods: Endogenous tryptic peptides were extracted from individual sepsis and control EDTA plasma samples in a step gradient of acetonitrile for random and independent sampling by LC-ESI-MS/MS with a set of robust and sensitive linear quadrupole ion traps. The MS/MS spectra were fit to fully tryptic peptides within proteins using the X!TANDEM algorithm. The protein observation frequency was counted using the SEQUEST algorithm after selecting the single best charge state and peptide sequence for each MS/MS spectra. The protein observation frequency of ICU-sepsis versus ICU Control was subsequently tested by Chi square analysis. The average protein or peptide log10 precursor intensity was compared across disease and control treatments by ANOVA in the R statistical system.Results: Peptides and/or phosphopeptides of common plasma proteins such as ITIH3, SAA2, SAA1, and FN1 showed increased observation frequency by Chi square (χ2 > 9, p < 0.003) and/or precursor intensity in sepsis. Cellular gene symbols with large Chi square values from tryptic peptides included POTEB, CTNNA1, U2SURP, KIF24, NLGN2, KSR1, GTF2H1, KIT, RPS6KL1, VAV2, HSPA7, SMC2, TCEB3B, ZNF300, SUPV3L1, ADAMTS20, LAMB4, MCCC1, SUPT6H, SCN9A, SBNO1, EPHA1, ABLIM2, cB5E3.2, EPHA10, GRIN2B, HIVEP2, CCL16, TKT, LRP2 and TMF1 amongst others showed increased observation frequency. Similarly, increased frequency of tryptic phosphopeptides were observed from POM121C, SCN8A, TMED8, NSUN7, SLX4, MADD, DNLZ, PDE3B, UTY, DEPDC7, MTX1, MYO1E, RXRB, SYDE1, FN1, PUS7L, FYCO1, USP26, ACAP2, AHI1, KSR2, LMAN1, ZNF280D and SLC8A2 amongst others. Increases in mean precursor intensity in peptides from common plasma proteins such as ITIH3, SAA2, SAA1, and FN1 as well as cellular proteins such as COL24A1, POTEB, KANK1, SDCBP2, DNAH11, ADAMTS7, MLLT1, TTC21A, TSHR, SLX4, MTCH1, and PUS7L among others were associated with sepsis. The processing of SAA1 included the cleavage of the terminal peptide D/PNHFRPAGLPEKY from the most hydrophilic point of SAA1 on the COOH side of the cystatin C binding that was most apparent in ICU-Sepsis patients compared to all other diseases and controls. Additional cleavage of SAA1 on the NH2 terminus side of the cystatin binding site were observed in ICU-Sepsis. Thus there was disease associated variation in the processing of SAA1 in ICU-Sepsis versus ICU controls or other diseases and controls.Conclusion: Specific proteins and peptides that vary between diseases might be discovered by the random and independent sampling of multiple disease and control plasma from different hospital and clinics by LC-ESI-MS/MS for storage in a relational SQL Server database and analysis with the R statistical system that will be a powerful tool for clinical research. The processing of SAA1 may play an unappreciated role in the inflammatory response to Sepsis.
Thavarajah T, Dos santos CC, Slutsky AS, et al. The plasma peptides of sepsis. Clin Proteomics. 2020;17:26.
Abstract: Infection, sterile injury, and chronic inflammation trigger the acute phase response in order to re-establish homeostasis. This response includes production of positive acute phase proteins in the liver, such as members of the serum amyloid A (SAA) family. In humans the major acute phase SAAs comprise a group of closely related variants of SAA1 and SAA2. SAA1 was proven to be chemotactic for several leukocyte subtypes through activation of the G protein-coupled receptor FPRL1/FPR2. Several other biological activities of SAA1, such as cytokine induction, reported to be mediated via TLRs, have been debated recently. Especially commercial SAA1, recombinantly produced in Escherichia coli, was found to be contaminated with bacterial products confounding biological assays performed with this rSAA1. We purified rSAA1 by RP-HPLC to homogeneity, removing contaminants such as lipopolysaccharides, lipoproteins and formylated peptides, and re-assessed several biological activities attributed to SAA1 (chemotaxis, cytokine induction, MMP-9 release, ROS generation, and macrophage differentiation). The homogeneous rSAA1 (hrSAA1) lacked most cell-activating properties, but its leukocyte-recruiting capacity in vivo and it's in vitro synergy with other leukocyte attractants remained preserved. Furthermore, hrSAA1 maintained the ability to promote monocyte survival. This indicates that pure hrSAA1 retains its potential to activate FPR2, whereas TLR-mediated effects seem to be related to traces of bacterial TLR ligands in the E. coli-produced human rSAA1
Abouelasrar salama S, De bondt M, De buck M, et al. Serum Amyloid A1 (SAA1) Revisited: Restricted Leukocyte-Activating Properties of Homogeneous SAA1. Front Immunol. 2020;11:843.
Abstract: Serum amyloid A (SAA), one of the major highly conserved acute-phase proteins in most mammals, is predominantly produced by hepatocytes and also by a variety of cells in extrahepatic tissues. It is well-known that the expression of SAA is sharply increased in bacterial infections. However, the exact physiological function of SAA during bacterial infection remains unclear. Herein, we showed that SAA expression significantly increased in abscesses of Staphylococcus aureus cutaneous infected mice, which exert direct antibacterial effects by binding to the bacterial cell surface and disrupting the cell membrane in acidic conditions. Mechanically, SAA disrupts anionic liposomes by spontaneously forming small vesicles or micelles under acidic conditions. Especially, the N-terminal region of SAA is necessary for membrane disruption and bactericidal activity. Furthermore, we found that mice deficient in SAA1/2 were more susceptible to infection by S. aureus In addition, the expression of SAA in infected skin was regulated by interleukin-6. Taken together, these findings support a key role of the SAA in host defense and may provide a novel therapeutic strategy for cutaneous bacterial infection.
Zheng H, Li H, Zhang J, et al. Serum amyloid A exhibits pH dependent antibacterial action and contributes to host defense against cutaneous infection. J Biol Chem. 2020;295(9):2570-2581.
Abstract: A natural leukocyte chemoattractant was isolated from bovine serum by an established 4-step purification procedure. Based on its relative molecular mass of 7287 and NH2-terminal sequence, the protein was identified as a carboxy-terminal peptide of the acute phase protein serum amyloid A1 (SAA1). This SAA1(46-112) fragment and its human equivalent SAA1(47-104) were chemically synthesized. Unlike intact SAA1α, these SAA fragments failed to directly chemoattract neutrophils and monocytes, to induce chemokines, and to stimulate downstream extracellular signal-regulated kinase signaling in monocytes. However, the SAA fragments potently synergized with CCL3 to induce monocyte migration and with CXCL8 to stimulate neutrophil shape changes and chemotaxis. Unlike intact SAA1α, SAA1(46-112) did not induce CXCL6 ex vivo but provoked a cooperative intraperitoneal neutrophil recruitment in mice when coinjected with CXCL6 into the peritoneal cavity. Moreover, SAA1(47-104) desensitized the synergy between intact SAA1α and CXCL8 in neutrophil chemotaxis, suggesting that this peptide binds formyl peptide receptor 2 (FPR2). This was evidenced by a complete blockade of synergy between the COOH-terminal SAA1 fragments and CXCL8 or CCL3 in neutrophil and monocyte chemotaxis, respectively, by the FPR2 antagonist WRW4 Thus, SAA1 is degraded into fragments lacking chemokine-inducing capacity, while keeping synergy with cytokine-induced chemokines to sustain limited inflammation.
De buck M, Gouwy M, Berghmans N, et al. COOH-terminal SAA1 peptides fail to induce chemokines but synergize with CXCL8 and CCL3 to recruit leukocytes via FPR2. Blood. 2018;131(4):439-449.
Abstract: Serum amyloid A1 (SAA1) is a prototypic acute phase protein, induced to extremely high levels by physical insults, including inflammation and infection. Human SAA and its NH2-terminal part have been studied extensively in the context of amyloidosis. By contrast, little is known about COOH-terminal fragments of SAA. Intact SAA1 chemoattracts leukocytes via the G protein-coupled receptor formyl peptide receptor like 1/formyl peptide receptor 2 (FPR2). In addition to direct leukocyte activation, SAA1 induces chemokine production by signaling through toll-like receptor 2. We recently discovered that these induced chemokines synergize with intact SAA1 to chemoattract leukocytes in vitro and in vivo. Gelatinase B or matrix metalloproteinase-9 (MMP-9) is also induced by SAA1 during infection and inflammation and processes many substrates in the immune system. We demonstrate here that MMP-9 rapidly cleaves SAA1 at a known consensus sequence that is also present in gelatins. Processing of SAA1 by MMP-9 at an accessible loop between two alpha helices yielded predominantly three COOH-terminal fragments: SAA1(52-104), SAA1(57-104), and SAA1(58-104), with a relative molecular mass of 5,884.4, 5,327.3, and 5,256.3, respectively. To investigate the effect of proteolytic processing on the biological activity of SAA1, we chemically synthesized the COOH-terminal SAA fragments SAA1(52-104) and SAA1(58-104) and the complementary NH2-terminal peptide SAA1(1-51). In contrast to intact SAA1, the synthesized SAA1 peptides did not induce interleukin-8/CXCL8 in monocytes or fibroblasts. Moreover, these fragments possessed no direct chemotactic activity for neutrophils, as observed for intact SAA1. However, comparable to intact SAA1, SAA1(58-104) cooperated with CXCL8 in neutrophil activation and migration, whereas SAA1(1-51) lacked this potentiating activity. This cooperative interaction between the COOH-terminal SAA1 fragment and CXCL8 in neutrophil chemotaxis was mediated by FPR2. Hence, proteolytic cleavage of SAA1 by MMP-9 fine tunes the inflammatory capacity of this acute phase protein in that only the synergistic interactions with chemokines remain to prolong the duration of inflammation.
Gouwy M, De buck M, Abouelasrar salama S, et al. Matrix Metalloproteinase-9-Generated COOH-, but Not NH-Terminal Fragments of Serum Amyloid A1 Retain Potentiating Activity in Neutrophil Migration to CXCL8, With Loss of Direct Chemotactic and Cytokine-Inducing Capacity. Front Immunol. 2018;9:1081.
Abstract: The proteins known as serum amyloid A (SAA) play major, but relatively uncharacterized, roles in the acute phase response and are important components of the innate immune system of humans and probably all vertebrates. N-terminal fragments of the inducible isoforms, SAA1 and SAA2, are the major constituents of fibrils formed during secondary or reactive amyloidosis. Little is known about the structure of SAA beyond secondary structure analyses and circular dichroism spectroscopic data indicating significant alpha helix conformation. Analysis of the primary structure of human SAA indicates probable homology to the N-terminal domain of hemocyanins of arthropods and suggests that approximately 80% of the molecule may consist of a helical bundle with the remaining portion of the C-terminus potentially disordered. This model of SAA suggests that proposed binding sites for laminin, fibronectin, and calcium are segregated to one face of the molecule and that the heparin/heparan binding site is found in the putatively disordered region of the protein. It is possible that removal of the N-terminal 76 amino acid fragment by proteolytic cleavage found generates an unstable entity that undergoes a helix to beta strand transition analogous to the fibril process of A-beta and prion peptides.
Stevens FJ. Hypothetical structure of human serum amyloid A protein. Amyloid. 2004;11(2):71-80.
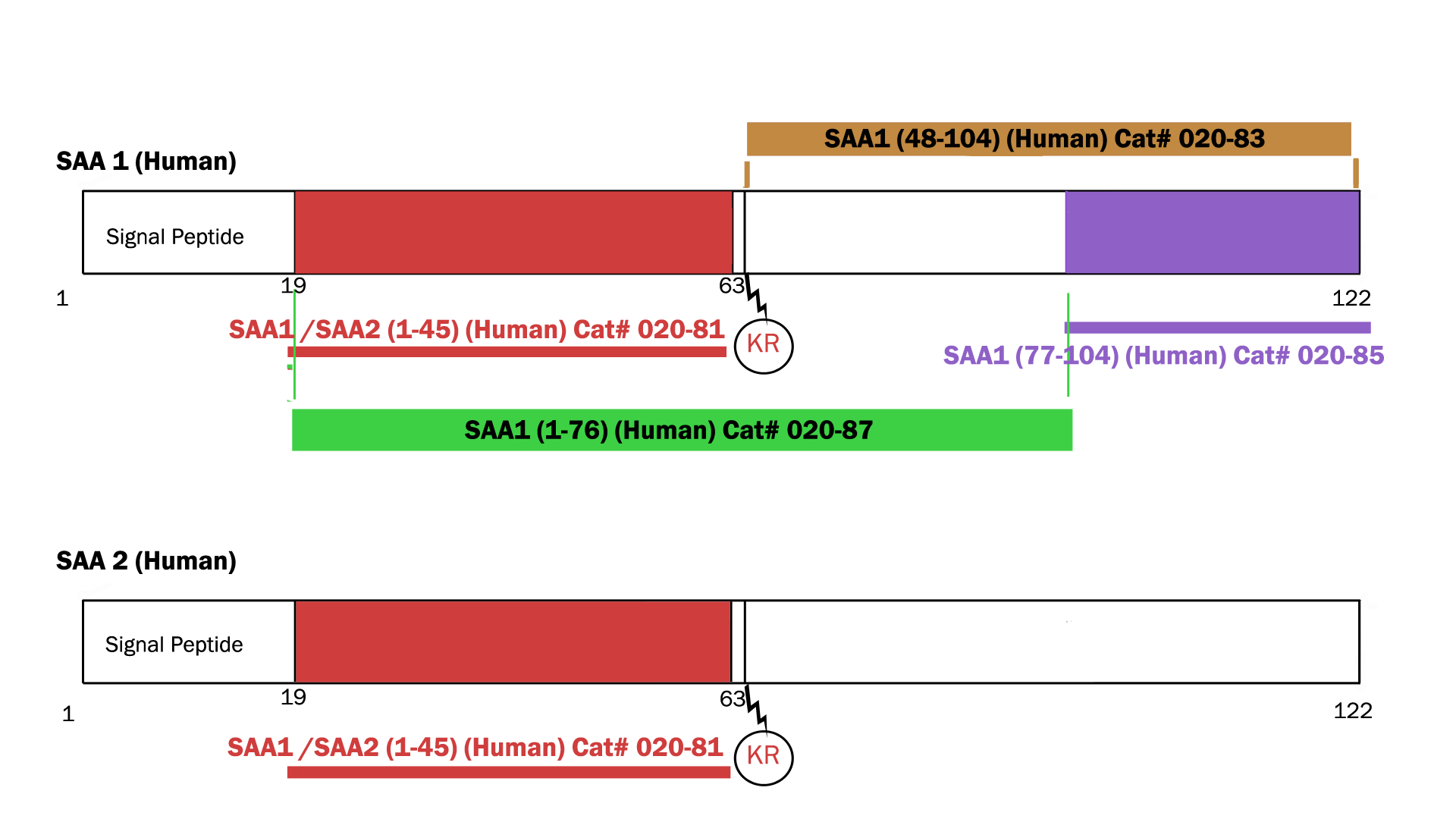
| Catalog# | Product | Standard Size | Price |
|---|---|---|---|
| EK-020-83 | SAA1(48-104) (Human) – EIA kit | 96 wells | $548 |
| 020-81 | SAA1(1-45) / SAA2 (1-45) (Human) | 100 µg | $415 |
| 020-84 | SAA1(47-104) (Human) | 100 µg | $457 |
| 020-83 | SAA1(48-104) (Human) | 100 µg | $450 |
| 020-85 | SAA1(77-104) (Human) | 100 µg | $356 |
| 020-87 | Synthetic SAA1 (1-76) protein (Human) | 100 µg | $505 |

Social Network Confirmation